Modèle conceptuel du simulateur
Dans cette partie, les différents modules de Syst'N sont présentés de manière détaillée
Module Croissance aérienne
Formalisme général
Ce module calcule deux variables de croissance des plantes : l’indice de surface foliaire (LAI = leaf area index) et la matière sèche aérienne. Tout d’abord, le LAI est calculé, de façon journalière, en fonction du stade de la culture, de la MSaérienne, de la teneur en azote critique, et d’un indice de stress azoté. Ensuite, ce module calcule la production de MSaérienne journalière : ∆MSaer(j) = xRUE(j) (paramètre de culture fonction du stade) xPAR(j) (calculé à partir du rayonnement global) xEi(j) (calculé à partir du LAI) xf’(T) (fonction température) xg’(H) (fonction de stress hydrique) xh(INN) (fonction de stress azoté) La MS aérienne totale du jour j est ensuite calculée en ajoutant la production de MS du jour à la MS totale du jour précédent : MSaer(j) = MSaer(j-1) + ∆MSaer(j) Enfin, la biomasse totale du couvert végétal est calculée à la fin de ce module en affectant un coefficient racinaire à la biomasse aérienne précédemment obtenue : MStotale(j) = MSaer(j) x COEF_RAC Avec MStotale(j) : quantité de matière sèche totale (parties aériennes + racinaires) au jour j (T.ha-1) COEF_RAC : coefficient racinaire de répartition de la matière sèche entre les racines et les parties aériennes propre à l’espèce
Formalisme de calcul de LAI
Si le stade LAImax n’est pas atteint alors : LAI(j)=PARAM_LAI×N_AER(j)×min(h(INN),TURFAC) Avec LAI : indice de surface foliaire (m2feuilles.m-2sol) PARAM_LAI : paramètre de croissance du LAI par unité d’azote assimilée par la plante (m2 de feuille.m-2 de sol.kgN-1) N_AER : quantité d’azote contenue dans les parties aériennes de la plante (kgN.ha-1) h(INN) : indice de stress azoté, si INN > 1 alors h(INN) = 1, si INN < 1 alors h(INN) = INN TURFAC : facteur de turgescence, i.e. stress hydrique appliqué au processus d’expansion foliaire (i.e. production de LAI) compris entre 0 et 1 (adimensionnel) Si le stade LAImax est atteint alors :
| LAI(j) = LAImax x | SOMTEMP_MP - SOMTEMP(j) |
| SOMTEMP_MP - SOMTEMP_LAIMAX |
Formalisme de calcul de RUE
Si le stade début remplissage des grains (DRG) n’est pas encore atteint alors : RUE(j) = RUE Avec RUE (Radiation Use Efficiency) : efficacité de conversion du rayonnement photosynthétiquement actif (PAR) intercepté en matière sèche, propre à chaque espèce (gMS.MJ-1)
 Le RUE correspond au coefficient Eb dans la formule de Monteith.
Le RUE correspond au coefficient Eb dans la formule de Monteith.
Si le stade DRG est atteint alors :
| RUE(j) = RUE x | SOMTEMP_MP - SOMTEMP(j) |
| SOMTEMP_MP - SOMTEMP_DRG |
Formalisme de calcul de PAR
Le rayonnement photosynthétiquement actif (PAR) est calculé comme suit : PAR(j) = c x RG(j) Avec c = 0.48, constante (Varlet-Grancher et al., 1982) RG(j) : rayonnement global incident le jour j (MJ.m-2)
Le rayonnement global incident est souvent compris entre 0.5 et 20 en période hivernale, et entre 2 et 50 en période estivale.
Formalisme de calcul de Ei
L’efficience d’interception du rayonnement par le couvert est calculé comme suit : Ei(j) = Eimax x (1 – e-k x LAI(j)) Avec Eimax : efficience maximale d’interception du rayonnement par le couvert (paramètre variant selon l’espèce) k : coefficient d’extinction du rayonnement dans le couvert (paramètre par espèce) LAI(j) : indice de surface foliaire au jour j (m2 de feuilles.m-2 de sol)
Formalisme de calcul de f'(T)
L’effet de la température sur l’efficacité de conversion du rayonnement en biomasse est modélisé selon le formalisme suivant : Si T < TEMPCULT_MIN alors f’(T)=0 Si TEMPCULT_MIN ≤ T < TEMPCULT_OPT alors
| f’(T)=1 - [ | T-TEMPCULT_OPT) | ]2 |
| TEMPCULT_MIN-TEMPCULT_OPT |
| f’(T)=1 - [ | T-TEMPCULT_OPT) | ]2 |
| TEMPCULT_MAX-TEMPCULT_OPT |
Formalisme de calcul du g'(H)
L’effet du stress hydrique sur la croissance aérienne et l'expansion foliaire est modélisé à partir de deux facsteurs de stress hydriques (issus du modèle STICS (Brisson et al,2009) : SWFAC, facteur de stress hydrique appliqué aux processus de photosynthèse (i.e. production de MS journalière) et de transpiration (i.e. absorption d’eau) et TURFAC, facteur de turgescence des cellules et est appliqué au processus d’expansion foliaire (i.e. production journalière de LAI). GH=min(1 ; SWFAC)
| SWFAC(j)= | SH(j-1) |
| BESOIN_EAU(j-1) |
| TURFAC(j) = | SWFAC(j) |
| 1.5 |
Formalisme de calcul du h(INN)
L’effet d’un stress azoté sur l’efficacité de conversion du rayonnement en biomasse est modélisé selon le formalisme suivant : h(INN)=min(1 ; INN) INN= %NA /%NC Avec INN : indice de nutrition azotée %NA : teneur en azote réelle des parties aériennes de la culture (%) %NC : teneur en azote critique des parties aériennes de la culture (%) (voir paragraphe suivant pour plus de détails)
Courbe de dilution critique de l’azote dans les parties aériennes
La teneur en azote critique se calcule comme suit : Si MSaer(j) < MSaer_seuil alors %NC = a x MSaer_seuil-b Si MSaer(j) ≥ MSaer_seuil alors %NC = a x MSaer(j)-b Avec MSaer(j) : quantité de matière sèche (t.ha-1) MSaer_seuil : quantité de matière sèche en-deçà de laquelle la courbe critique décrit un plateau (quantités de matière sèche faibles)(T.ha-1) %NC : teneur en azote critique des parties aériennes de la culture (%) a, b : paramètres de la courbe critique, connus par espèce
La courbe critique a été validée jusqu’à floraison, en général, pour les différentes espèces. Après ce stade, plusieurs travaux montrent, soit que la courbe initiale peut être maintenue, soit que la teneur en azote critique des parties aériennes de la culture reste stable. Ici, on propose de garder la même courbe critique.
Et pour les prairies
La croissance en biomasse de la prairie repose sur des valeurs de croissance potentielle maximale journalières, affectées par un facteur de stress azoté, un facteur de stress hydrique, un facteur température et un facteur biomasse. Le modèle est de forme multiplicative :
∆MSaer(j) = CJS(j) x fbiom(MS) x k1(stressN) x k2(stressH) x k3(T°)
Avec
∆MSaer(j) : croissance journalière de matière sèche (kgMS.ha-1)
CJS (j) : croissance journalière standard (kgMS.ha-1)
k1(stressN), k2(stressH), k3(T°) : stress azoté, hydrique, température, respectivement
Fbiom : fonction de biomasse
La croissance journalière standard correspond à des valeurs de croissance potentielle de MS définies selon le jour julien considéré. Les valeurs sont représentées dans la figure 9.
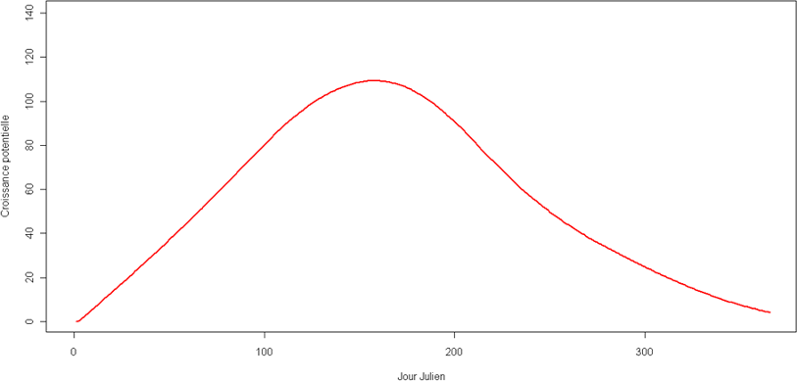 Figure 9 : Courbe de croissance journalière standard pour les prairies
L’indice de stress azoté k1 est fonction une fonction linéaire de l’indice de nutrition azoté INN, présenté précedemment.
L’indice de stress hydrique k2 est une fonction linéaire de Gh, présentée précedemment.
L'indice de stress températude k3 provient d'une fonction non liénaire dépendant de la température. La courbe est présentée en figure 10.
Figure 9 : Courbe de croissance journalière standard pour les prairies
L’indice de stress azoté k1 est fonction une fonction linéaire de l’indice de nutrition azoté INN, présenté précedemment.
L’indice de stress hydrique k2 est une fonction linéaire de Gh, présentée précedemment.
L'indice de stress températude k3 provient d'une fonction non liénaire dépendant de la température. La courbe est présentée en figure 10.
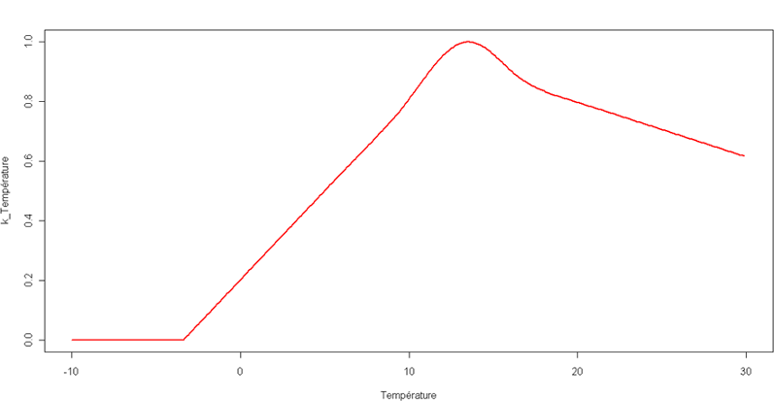 Figure 10 : Courbe de la fonction de stress température pour les prairies
Figure 10 : Courbe de la fonction de stress température pour les prairies
Cas particulier des cultures présentant une période de dormance hivernale
Dans ce cas, la croissance est inhibée lorsque la température descend en-dessous d'un certain seuil (spécifique à chaque culture) pendant plus de 7 jours et une fois la date de début potentiel de dormance dépassée (date également dépendant de la culture considérée). De la même manière la croissance reprend, une fois la date potentielle de fin de dormance passée et après 7 jours au-dessus de la température seuil.
